160 млн, лет назад богатый растительный мир обеспечивал пищей возникших к этому времени гигантских зауропод, а также давал укрытие огромному числу мелких млекопитающих и ящеров. В это время были широко распространены хвойные, папоротники, хвощи, древовидные папоротники и саговники.
Отличительной чертой юрского периода было появление и расцвет гигантских ящеротазовых травоядных динозавров, зауропод, самых крупных из когда-либо существовавших наземных животных. Несмотря на свои размеры, эти динозавры были достаточно многочисленными.Их окаменевшие останки находят на всех континентах (за исключением Антарктиды) в породах от раннего юрского до позднего мелового периодов , хотя самыми распространенными они были во второй половине юрского периода. В это же время зауроподы достигают своих самых больших размеров. Они просуществовали до позднего мелового периода , когда среди наземных травоядных стали доминировать огромные гадрозавры («утконосые динозавры»).
Внешне все зауроподы выглядели похожими друг на друга: с чрезвычайно длинной шеей, еще более длинным хвостом, массивным, но относительно коротким туловищем, четырьмя колонноподобными ногами и сравнительно небольшой головой. У различных видов могли изменяться только положение тела и пропорции отдельных частей. Например, такие зауроподы позднеюрского периода, как брахиозавры (Brachiosaurus - «плечистый ящер»), в плечевом поясе были выше, чем в тазовом, в то время как современные им диплодоки (Diplodocus - «двойной отросток») были значительно ниже, и при этом их бедра возвышались над плечами. У некоторых видов зауропод, таких как камаразавры (Camarasaurus - «камерный ящер»), шея была относительно короткой, только немного длиннее тела, а у других, например диплодоков , она была длиннее тела более чем в два раза.
Зубы и способ питания
Внешнее сходство зауропод маскирует неожиданно широкое разнообразие строения их зубов и, следовательно, методов питания.
Череп диплодока помог палеонтологам понять способ питания этого динозавра. Истирание зубов свидетельствует, что он срывал листья либо снизу, либо сверху над собой.
Раньше во многих книгах о динозаврах упоминались «мелкие, тонкие зубы» зауропод , но сейчас известно, что зубы некоторых из них, например камаразавров, были достаточно массивными и прочными, чтобы измельчать даже очень твердую растительную пищу, в то время как длинные и тонкие, напоминающие карандаши зубы диплодока действительно выглядят неспособными выдерживать значительные нагрузки, возникающие при пережевывании твердых растений.

диплодок (Diplodocus). Длинная шея позволяла ему «счесывать» пищу с самых высоких хвойных растений. Считается, что диплодоки жили небольшими стаданил и питались побегами деревьев.
При исследовании зубов диплодоков , проведенном в последние годы в Англии, была обнаружена необычная изношенность их боковых поверхностей. Такой характер истирания зубов дал ключ к пониманию того, каким образом могли питаться эти огромные животные. Боковая поверхность зубов могла истереться, только если между ними что-то двигалось. По-видимому, диплодок пользовался своими зубами для раздирания пучков листьев и побегов, действуя ими как гребнем, при этом его нижняя челюсть могла немного сдвигаться вперед и назад. Скорее всего, когда животное разделяло на полоски растения, захваченные внизу, двигая головой вверх и назад, нижняя челюсть была смещена назад (верхние зубы располагались перед нижними), а когда оно тянуло расположенные вверху ветви высоких деревьев вниз и назад, то выдвигало нижнюю челюсть вперед (нижние зубы оказывались перед верхними).
Брахиозавр , вероятно, использовал свои более короткие, слегка заостренные зубы для срывания только высоко расположенных листьев и побегов, поскольку ориентация его тела в вертикальном направлении, обусловленная большей длиной передних ног, затрудняла питание растениями, растущими невысоко над почвой.
Узкая специализация
Камаразавр, размерами несколько уступающий упомянутым выше гигантам, обладал относительно короткой и более толстой шеей и, скорее всего, питался листьями, расположенными на промежуточной высоте между уровнями питания брахиозавров и диплодоков . У него был высокий, закругленный и более массивный, по сравнению с другими зауроподами , череп, а также более массивная и прочная нижняя челюсть, что свидетельствует о лучшей способности измельчать твердую растительную пищу.Описанные выше детали анатомического строения зауропод показывают, что в пределах одной экологической системы (в лесах, покрывающих в это время большую часть суши) зауроподы питались различной растительной пищей, по-разному добывая ее на различных уровнях. Это разделение по стратегии питания и по виду пищи, которое можно видеть в сообществах травоядных животных и сегодня, получило название «тропического секционирования».

Брахиозавры (Brachiosaurus) достигали более 25 м в длину и 13 м в высоту. Их окаменевшие останки и окаменевшие яйца находят в Восточной Африке и Северной Америке. Вероятно, они жили стадами подобно современным слонам.
Основное отличие сегодняшних экосистем травоядных животных от экосистем позднего юрского периода, в которых доминировали зауроподы , касается только массы и высоты животных. Ни одно из современных травоядных животных, включая слонов и жирафов, не достигает высоты, сравнимой с высотой большинства крупных зауропод, и ни одному из современных сухопутных животных не требуется такое огромное количество пищи, как этим гигантам.
Другой край шкалы
Некоторые жившие в юрском периоде зауроподы достигали фантастических размеров, например, напоминающий брахиозавра суперзавр (Supersaurus), чьи останки найдены в США (штат Колорадо) весил, вероятно, около 130 т, т. е. был во много раз больше крупного самца африканского слона. Но эти сверхгиганты делили сушу с прячущимися под землей крохотными существами, не принадлежащими к динозаврам и даже к рептилиям. Юрский период был временем существования много численных древних млекопитающих. Эти мелкие, покрытые мехом, живородящие и выкармливающие детенышей молоком теплокровные животные получили название многобугорчатых из-за необычного строения своих коренных зубов: многочисленные, сросшиеся вместе цилиндрические «бугорки» образовывали неровные поверхности, великолепно приспособленные к измельчению растительной пищи.Многобугорчатые были самой многочисленной и самой разнообразной группой млекопитающих юрского и мелового периодов . Это единственные всеядные млекопитающие мезозойской эры (остальные были специализированными насекомоядными или плотоядными). Они известны с позднеюрских отложений, но последние находки показывают, что они близки к малоизвестной группе чрезвычайно древних млекопитающих позднего триаса, т. н. харамиидам.
Строением черепа и зубов многобугорчатые очень напоминали сегодняшних грызунов, у них было две пары выдающихся вперед резцов, придававших им вид типичного грызуна. За резцами находился не содержавший зубов промежуток, за которым до самого конца маленьких челюстей шли коренные зубы. Однако ближайшие к резцам зубы многобугорчатых имели необычную структуру. В действительности это были первые ложнокоренные (премолярные) зубы с искривленными пилообразными краями.
Такое необычное строение зубов в процессе эволюции повторно возникло у некоторых из современных сумчатых, например, у крысиных кенгуру в Австралии, зубы которых такой же формы и находятся в таком же месте челюсти, как и ложнокоренные зубы многобугорчатых. При пережевывании пищи в момент смыкания челюстей многобугорчатые могли смещать нижнюю челюсть назад, перемещая эти острые пилообразные зубы поперек пищевых волокон, а длинные резцы могли использоваться для прокалывания плотных растений или твердых наружных скелетов насекомых.

Ящеротазовый мегалозавр (Megalosaurus) и его детеныши, настигшие птицетазового сцелидозавра (Scelidosaurus). Сцелидозавр - древний вид динозавров юрского периода с неравномерно развитыми конечностями, достигавший 4 м в длину. Его спинной панцирь помогал защищаться от хищников.
Комбинация острых передних резцов, зазубренных лезвий и жевательных зубов означает, что питательный аппарат многобугорчатых был достаточно универсальным. Сегодняшние грызуны тоже очень успешная группа животных, процветающая в самых различных экологических системах и местах обитания. Скорее всего, именно высокоразвитый зубной аппарат, позволяющий питаться различной пищей, и стал причиной эволюционного успеха многобугорчатых. Их окаменевшие останки, встречающиеся на большинстве континентов, принадлежат различным видам: одни из них, по-видимому, жили на деревьях, в то время как другие, напоминающие современных песчанок, вероятно, были приспособлены к существованию в засушливом пустынном климате.
Изменение экосистем
Существование многобугорчатых охватывает период 215 млн. лет, простираясь от позднего триаса через всю мезозойскую эру до олигоценовой эпохи кайнозойской эры. Этот феноменальный успех, уникальный для млекопитающих и большинства наземных тетрапод, делает многобугорчатых самой успешной группой млекопитающих.Экосистемы мелких животных юрского периода включали также небольших ящериц самых различных видов и даже их водные формы.

Thrinadoxon (вид цинодонт). Его конечности слегка выдавались в стороны, а не располагались под туловищем, как у современных млекопитающих.
Они и дожившие до этого времени редко встречающиеся рептилии группы синапсид («зверообразных рептилий»), тритилодонты, жили в одно время и в тех же экосистемах, что и многобугорчатые млекопитающие. Тритилодонты были многочисленным и широко распространенным видом на протяжении всего триасового периода, но, подобно другим цинодонтам, очень пострадали во время позднетриасового вымирания. Это единственная группа цинодонтов, сохранившаяся в юрском периоде. По внешнему виду они, как и многобугорчатые млекопитающие, очень напоминали современных грызунов. Т. е. существенная часть экосистем мелких животных юрского периода состояла из животных, напоминающих грызунов: трилодонтов и многобугорчатых млекопитающих.
Многобугорчатые млекопитающие были, безусловно, наиболее многочисленной и разнообразной группой млекопитающих юрского периода, но в это время существовали и другие группы млекопитающих, включая: морганакодонтов (древнейших млекопитающих), амфилестид (amphilestids), пермурид (peramurids), амфитерид (amphitherids), тинодонтов (tinodontids) и докодонтов (docodonts). Все эти мелкие млекопитающие выглядели подобно мышам или землеройкам. У докодонтов, например, развивались своеобразные, широкие коренные зубы, хорошо приспособленные для пережевывания твердых семян и орехов.
В конце юрского периода существенные изменения происходили и на другом конце размерной шкалы в группе крупных двуногих хищных динозавров, теропод, представленных в это время аллозаврами (AUosaurus - «странные ящеры»). В конце юрского периода обособилась группа теропод, получившая название спинозаурид («колючие или шипастые ящеры»), отличительной чертой которых был гребень из длинных отростков туловищных позвонков, который, возможно, как спинной парус у некоторых пеликозавров, помогал им регулировать температуру тела. Такие спинозауриды , как Siamosaurus («ящер из Сиама»), длина которых достигала 12 м, вместе с другими тероподами делили в экосистемах того времени нишу крупнейших хищников.
У спинозаурид были незазубренные зубы и удлиненные, менее массивные по сравнению с другими тероподами этого времени, черепа. Эти особенности строения свидетельствуют, что способом питания они отличались от таких теропод, как аллозавры , Eustreptospondylus («сильно изогнутые позвонки») и цератозавры (Ceratosaurus - «рогатый ящер»), и, скорее всего, охотились на другую добычу.
Птицеподобные динозавры
В позднеюрское время возникли и другие виды теропод, очень отличавшиеся от таких огромных, массой до 4 тонн, хищников, как аллозавры . Это были орнитоминиды - длинноногие, длинношеие, с небольшими головами, беззубые всеядные животные, поразительно напоминающие современных страусов , отчего и получили свое название «имитаторы птиц».Самая первая орнитоминида, Elaphrosaums («легкий ящер»), из позднеюрских отложений Северной Америки имела легкие, пустотелые кости и беззубый клюв, а ее конечности, как задние, так и передние, были короче, чем у более поздних орнитоминид мелового периода , и, соответственно, это было более медлительное животное.
Другая экологически важная группа динозавров, возникшая в позднем юрском периоде, - нодозавры, четвероногие динозавры с массивными, покрытыми панцирями телами, короткими, относительно тонкими конечностями, узкой головой с удлиненной мордой (но с массивными челюстями), с небольшими листовидными зубами и роговым клювом. Их название («узловатые ящеры») связано с покрывающими кожу костяными пластинами, выступающими отростками позвонков и разбросанными по коже наростами, которые служили защитой от нападения хищников. Широкое распространение нодозавры получили только в меловом периоде , а в позднеюрское время они наряду с огромными, питающимися побегами деревьев зауроподами были только одним из элементов сообщества травоядных динозавров, служивших добычей целому ряду огромных хищников.
Юрский геологический период, Юра, Юрская система, средний период мезозоя. Начался 200-199 млн л. н. и окончился 144 млн. л. н.
Впервые отложения данного периода были обнаружены и описаны в Юре (горы в Швейцарии и Франции), отсюда и произошло название периода. Отложения юрского периода очень разнообразны: известняки, обломочные породы, сланцы, магматические породы, глины, пески, конгломераты, сформировавшиеся в разнообразнейших условиях. Отложения того времени довольно разнообразны: известняки, обломочные породы, сланцы, магматические породы, глины, пески, конгломераты, сформировавшиеся в разнообразнейших условиях.
Тектоника юрского периода: В начале юрского периода единый суперконтинент Пангея начал распадаться на отдельные континентальные блоки. Между ними образовывались мелководные моря. Интенсивные тектонические движения в конце триасового и в начале юрского периодов способствовали углублению больших заливов, постепенно отделивших Африку и Австралию от Гондваны. Углубился залив между Африкой и Америкой. В Евразии сформировались впадины: Немецкая, Англо-Парижская, Западно-Сибирская. Арктическое море залило северное побережье Лавразии. Именно благодаря этому климат юрского периода и стал более влажным. В юрский период начинают формироваться очертания материков: Африка, Австралия, Антарктида, Северная и Южная америки. И хотя они расположены иначе чем сейчас, но образовались они именно в юрский период.
Климат и растительность юрского периода
Вулканическая активность конца триасового - начала юрского периода вызвала трансгрессию моря. Материки разделились и климат в юрский период стал более влажным чем в триасовый. На месте пустынь триасового периода, в юрский период разрослась буйная растительность. Огромные территории покрылись пышной растительностью. Леса юрского периода в основном состояли из папоротников и голосеменных.
Теплый и влажный климат юрского периода способствовал буйному развитию растительного мира планеты.
Папоротникообразные, хвойные и цикадовые образовывали обширные болотистые леса. На побережье произрастали араукарии, туи, цикадовые. Папоротники и хвощи образовывали обширные лесные массивы. В начале юрского периода, около 195 млн. л. н. на всей территории северного полушария растительность была довольно однообразной. В северном растительном поясе преобладали гинкговые и травянистые папоротники. В юрский период гинкговые были распространены очень широко. Рощи гинкговых деревьев росли по всему поясу.
В южном растительном поясе преобладали цикадовые и древовидные папоротники.
Папоротники юрского периода и в наши дни сохранились в некоторых уголках дикой природы. Хвощи и плауны почти не отличались от современных.
животные: Юрский период - рассвет эпохи динозавров. Именно буйное развитие растительности способствовало появлению множества видов растительноядных динозавров. Рост численности растительноядных динозавров дал толчок к росту численности хищников. Динозавры расселились по всей суше и жили в лесах, озерах, болотах. Диапазон различий между ними настолько велик, что родственные связи между ними устанавливаются с большим трудом. Разнообразие видов динозавров в юрский период было велико. Они могли быть величиной с кошку или курицу, а могли достигать размеров огромных китов.
Юрский период - время обитания многих известных динозавров. Из ящеротазовых это аллозавр и диплодок. Из птицетазовых это стегозавр.
В юрский период в воздухе безраздельно царствовали крылатые ящеры - птерозавры. Они появились еще в триасе, но их расцвет приходился именного на юрский период Птерозавры были представлены двумя группами птеродактили и рамфоринхи.
В юрский период появляются первые птицы или нечто среднее между птицами и ящерами. Существа, появившиеся в юрский период и обладающие свойствами ящеров и современных птиц называются археоптериксы. Первые птицы - археоптериксы, величиной, с голубя. Археоптериксы жили в лесах. Питались преимущественно насекомыми и семенами.
Двустворчатые моллюски вытесняют с мелководий брахиопод. Брахиоподовые ракушечники замещаются устричными. Двустворчатые моллюски заполняют все жизненные ниши морского дна. Многие перестают собирать пищу с грунта и переходят к прокачиванию воды с помощью жабр. В теплых и мелких морях юрского периода происходили и другие важные события.
Юрский период породил множество видов плезиозавров и ихтиозавров, соперничавших с быстроходными акулами и чрезвычайно подвижными костными рыбами. а в морских глубинах леоплеврадон без остановки патрулировал свою территорию в поисках пищи.
Но одно существо по праву можно было назвать хозяином юрских морей. Это гигантский лиоплевродон весом до 25 тонн. Лиоплевродон был самым опасным хищником морей юрского периода, а возможно и за всю историю планеты.
Ю́рский пери́од (юра́) - средний (второй) период мезозойской эры. Начался 201,3 ± 0,2 млн лет назад, закончился 145,0 млн лет назад. Продолжался, таким образом, около 56 млн лет. Комплекс отложений (горных пород), соответствующих данному возрасту, называется ю́рской систе́мой. В разных регионах планеты эти отложения различаются по составу, генезису, внешнему виду.
Впервые отложения данного периода были описаны в Юре (горы в Швейцарии и Франции); отсюда и произошло название периода. Отложения того времени довольно разнообразны: известняки, обломочные породы, сланцы, магматические породы, глины, пески, конгломераты, сформировавшиеся в разнообразнейших условиях.
Флора
В юре огромные территории покрылись пышной растительностью, прежде всего разнообразными лесами. Они в основном состояли из папоротников и голосеменных.
Саговники - класс голосеменных, преобладавший в зелёном покрове Земли. Ныне они встречаются в тропиках и субтропиках. Под тенью этих деревьев бродили динозавры. Внешне саговники настолько охожи на невысокие (до 10-18 м) пальмы, что даже Карл Линней поместил их в своей системе растений среди пальм.
В юрский период рощи гингковых деревьев росли по всему тогдашнему умеренному поясу. Гинкго - листопадные (что необычно для голосеменных) деревья с кроной как у дуба и с небольшими веерообразными листьями. До наших дней сохранился только один вид - гинкго двулопастный.
Очень разнообразны были хвойные, похожие на современные сосны и кипарисы, которые процветали в то время не только в тропиках, но уже освоили и умеренный пояс. Папоротники постепенно исчезали.
Фауна
Морские организмы
По сравнению с триасом очень изменилось население морского дна. Двустворчатые моллюски вытесняют с мелководий брахиопод. Брахиоподовые ракушечники замещаются устричными. Двустворчатые моллюски заполняют все жизненные ниши морского дна. Многие перестают собирать пищу с грунта и переходят к прокачиванию воды с помощью жабер. Складывается новый тип рифовых сообществ, примерно такой же, как существует сейчас. Его основу составляют появившиеся в триасе шестилучевые кораллы.
Наземные животные Юрского периода
Одним из ископаемых существ, сочетающих признаки птиц и рептилий, является археоптерикс, или первоптица. Впервые его скелет обнаружили в так называемых литографских сланцах в Германии. Находка была сделана через два года после выхода труда Чарльза Дарвина «Происхождение видов» и стала веским аргументом в пользу теории эволюции. Летал археоптерикс ещё довольно плохо (планировал с дерева на дерево), а размером был примерно с ворону. Вместо клюва он имел пару зубастых, хотя и слабых челюстей. На крыльях его были свободные пальцы (из современных птиц они сохранились лишь у птенцов гоацинов).
В юрский период на Земле обитают мелкие, покрытые шерстью теплокровные животные - млекопитающие. Они живут рядом с динозаврами и почти незаметны на их фоне. В юре произошло разделение млекопитающих на однопроходных, сумчатых и плацентарных.
Динозавры (англ. Dinosauria, от др.-греч. δεινός - страшный, ужасный, опасный и σαύρα - ящер, ящерица) обитали в лесах, озёрах, болотах. Диапазон различий между ними настолько велик, что родственные связи между ними устанавливаются с большим трудом. Встречались динозавры с размерами от кошки до кита. Разные виды динозавров могли передвигаться на двух или четырёх конечностях. Среди них были как хищники, так и травоядные животные.
Шкала
| Геохронологическая шкала | |||
|---|---|---|---|
| Эон | Эра | Период | |
| Ф а н е р о з о й | Кайнозой | Четвертичный | |
| Неоген | |||
| Палеоген | |||
| Мезозой | Мел | ||
| Юра | |||
| Триас | |||
| Палеозой | Пермь | ||
| Карбон | |||
| Девон | |||
| Силур | |||
| Ордовик | |||
| Кембрий | |||
| Д о к е м б р и й | П р о т е р о з о й | Нео- протерозой | Эдиакарий |
| Криогений | |||
| Тоний | |||
| Мезо- протерозой | Стений | ||
| Эктазий | |||
| Калимий | |||
| Палео- протерозой | Статерий | ||
| Орозирий | |||
| Риасий | |||
| Сидерий | |||
| А р х е й | Неоархей | ||
| Мезоархей | |||
| Палеоархей | |||
| Эоархей | |||
| Катархей | |||
Подразделение юрской системы
Юрская система подразделяется на 3 отдела и 11 ярусов:
| система | отдел | ярус | Возраст, млн лет назад | |
|---|---|---|---|---|
| Мел | Нижний | Берриасский | меньше | |
| Юрский период | Верхняя (мальм) | Титонский | 145,0-152,1 | |
| Киммериджский | 152,1-157,3 | |||
| Оксфордский | 157,3-163,5 | |||
| Средняя (доггер) | Келловейский | 163,5-166,1 | ||
| Батский | 166,1-168,3 | |||
| Байосский | 168,3-170,3 | |||
| Ааленский | 170,3-174,1 | |||
| Нижняя (лейас) | Тоарский | 174,1-182,7 | ||
| Плинсбахский | 182,7-190,8 | |||
| Синемюрский | 190,8-199,3 | |||
| Геттангский | 199,3-201,3 | |||
| Триас | Верхний | Рэтский | больше | |
| Подразделы даны в соответствии с IUGS по состоянию на январь 2013 | ||||

Ростры белемнитов Acrofeuthis sp. Ранний мел, готерив
Раковины брахиопод Kabanoviella sp. Ранний мел, готерив
Раковина двустворчатого моллюска Inoceramus aucella Trautschold, Ранний мел, готерив

Скелет морского крокодила стенеозавра, Steneosaurus boltensis Jaeger. Ранняя юра, Германия, Гольцмаден. Среди морских крокодилов - талаттозухий стенеозавр был наименее специализированной формой. У него были развиты не ласты, а обычные пятипалые конечности, как у наземных животных, хотя и несколько укороченные. Кроме того, на спине и брюхе сохранился мощный костный панцирь из пластин.
Три из представленных на стене образцов (крокодил стенеозавр и два ихтиозавра - стеноптеригий и эвринозавр) были найдены на одном из крупнейших в мире местонахождений раннеюрской морской фауны ГОЛЬЦМАДЕН (около 200 млн. лет назад; Бавария, Германия). Несколько столетий здесь велась разработка сланцев, использовавшихся как строительный и декоративный материал.
При этом было обнаружено огромное количество остатков беспозвоночных рыб, ихтиозавров, плезиозавров и крокодилов. Только скелетов ихтиозавров добыто более 300.


Небольшие летающие ящеры - сордесы были многочисленны в окрестностях озера Каратау. Питались они, вероятно, рыбой и насекомыми. На некоторых экземплярах сордесов сохранились остатки волосяного покрова, что крайне редко встречается в других местонахождениях.


Текодонты - группа предновая для остальных архозавров. Первые представители (1,2) были наземными хищниками с широко расставленными конечностями. В процессе эволюции одни текодонты приобрели полувертикальную и вертикальную постановку лап при четвероногом способе передвижения (3,5,6), другие - паралельно сразвитием двуногости (2,7,8). Большинство текодонтов было наземными, но часть из них вела амфибиотический образ жизни (6).

Крокодилы близки к текодонтам. Ранние крокодилы (1,2,9) были наземными животными, в мезозое также существовали морские формы с ластами и хвостовым плавником (10), а современные крокодилы приспособлены к амфибиотическому образу жизни (11).
Динозавры - центральная и наиболее яркая группа архозавров. Крупные хищные карнозавры (14,15) и мелкие хищные цепурозавры (16,17,18), а также растительноядные орнитоподы (19,20,21,22) были двуногими. Другие использовали четвероногий способ передвижения: завроподы (12,13), цератопсы (23), стегозавры(24) и антипозавры (25). Завроподы и утконосые динозавры (21) в той или иной степени перешли к амфибиотическому образу жизни. Одним из наиболее высокоорганизованных среди архозавров были летающие ящеры (26,27,28), имевшие крылья с летательной перепонкой, волосяной покров и, возможно, постоянную температуру тела.
Птицы
- считаются непосредственными потомками мезозойских архозавров.

Мелкие наземные крокодилы, объединяемые в группу нотозухий (Notosuchia), были широко распространены в Африке и Южной Америке на протяжении мелового периода.

Часть черепа морского ящера - плиозавра. Pliosaurus cf. grandis Owen, Поздняя юра, Поволжье. Плиозавры, также как и их ближайшие родственники - плезиозавры, были прекрасно адаптированы к водной среде. Отличались крупной головой, короткой шеей и длинными мощными ластовидными конечностями. Большинство плиозавров обладало кинжаловидными зубами, и они были самыми опасными хищниками морей юрского периода. Этот образец, длиной 70 см - лишь передняя треть черепа плиозавра, а полная длина животного составляла 11-13 м. плиозавр жил 150-147 млн. лет назад.


Личинка жука-коптоклавы, Coptoclava longipoda Ping. Это один из самых опасных хищников в озере.
По-видимому, в середине мелового периода условия в озерах сильно изменились и многим беспозвоночным пришлось уйти в реки, ручьи или временные водоемы(ручейники, личинки которых строят домики-трубочки из песчинок; вислокрылки, двустворчатые рачки). Донные осадки этих водоемов не сохраняются, текучие воды размывают их, разрушая остатки животных и растений. Организмы, ушедшие в такие места обитания, исчезают из палеонтологической летописи.
Домики из песчинок, которые строили и носили на себе личинки ручейников, очень характерны для раннемеловых озер. В более поздние эпохи такие домики встречаются, в основном, в текучих водах
Личинки ручейника Terrindusia (реконструкция)

От: , 8624 просмотров
И Швейцарии. Начало юрского периода радиометрическим методом определяется в 185±5 млн. лет, конец — в 132±5 млн. лет; общая продолжительность периода около 53 млн. лет (по данным 1975).
Юрская система в её современном объёме выделена в 1822 немецким учёным А. Гумбольдтом под названием "юрская формация" в горах Юра (Швейцария), Швабский и Франконский Альб (). На территории юрские отложения впервые были установлены немецким геологом Л. Бухом (1840). Первая схема их стратиграфии , расчленения разработана русским геологом К. Ф. Рулье (1845-49) в Подмосковье.
Подразделения . Все основные подразделения юрской системы, впоследствии вошедшие в общую стратиграфическую шкалу, выделены на территории Центральной Европы и Великобритании . Расчленение юрской системы на отделы предложено Л. Бухом (1836). Основы ярусного расчленения юры заложены французским геологом А. д"Орбиньи (1850-52). Немецкий геолог А. Оппель впервые произвел (1856-58) детальное (зональное) подразделение юрских отложений. См. табл.
Большинство зарубежных геологов относит келловейский ярус к среднему отделу, мотивируя это приоритетом трёхчленного деления юры (чёрная, бурая, белая) Л. Буха (1839). Титонский ярус выделяется в отложениях Средиземноморской биогеографической провинции (Оппель, 1865); для северной (бореальной) провинции его эквивалентом является волжский ярус, впервые выделенный в Поволжье (Никитин, 1881).
Общая характеристика . Юрские отложения широко распространены на территории всех континентов и присутствуют в периферии, частях океанских впадин, слагая основание их осадочного слоя. К началу юрского периода в структуре земной коры обособляются два крупных континентальных массива: Лавразия , включавшая в себя платформы и палеозойские складчатые области Северной Америки и Евразии , и Гондвана , объединявшая платформы Южного полушария. Они разделялись Средиземноморским геосинклинальным поясом , который представлял собой океанический бассейн Тетис . Противоположное полушарие Земли занимала впадина Тихого океана , по окраинам которой развивались геосинклинальные области Тихоокеанского геосинклинального пояса .
В океаническом бассейне Тетис в течение всего юрского периода происходило накопление глубоководных кремнистых, глинистых и карбонатных отложений, сопровождавшееся местами проявлениями подводного толеит-базальтового вулканизма. Широкая южная пассивная окраина Тетиса являлась областью накопления мелководных карбонатных отложений. На северной окраине, которая в разных местах и в разное время имела как активный, так и пассивный характер, состав отложений более пёстрый: песчано-глинистые, карбонатные, местами флишевые, иногда с проявлением известково-щелочного вулканизма. Геосинклинальные области Тихоокеанского пояса развивались в режиме активных окраин. В них резко преобладают песчано-глинистые отложения, много кремнистых, очень активно проявлялась вулканическая деятельность. Основная часть Лавразии в ранней и средней юре представляла собой сушу. Морские трансгрессии из геосинклинальных поясов захватывали в ранней юре лишь территории Западной Европы, северную часть Западной Сибири, восточную окраину Сибирской платформы , а в средней юре и южную часть Восточно-Европейской. В начале поздней юры трансгрессия достигла максимума, распространившись на западную часть Североамериканской платформы, восточно-Европейской, всю Западную Сибирь, Предкавказье и Закаспий. Гондвана оставалась сушей на протяжении всего юрского периода. Морские трансгрессии с южной окраины Тетиса захватывали только северо-восточную часть Африканской и северо-западную часть Индостанской платформ . Моря в пределах Лавразии и Гондваны представляли собой обширные, но мелководные эпиконтинентальные бассейны, где накапливались маломощные песчано-глинистые, а в поздней юре в областях, прилегающих к Тетису, — карбонатные и лагунные (гипсо- и соленосные) отложения. На остальной территории юрские отложения либо отсутствуют, либо представлены континентальными песчано-глинистыми, часто угленосными толщами , заполняющими отдельные впадины. Тихий океан в юрском периоде представлял собой типичную океаническую впадину, где накапливались маломощные карбонатно-кремнистые осадки и покровы толеитовых базальтов , сохранившиеся в западной части впадины. В конце средней — начале поздней юры начинается формирование "молодых" океанов; происходит раскрытие Центральной Атлантики, Сомалийской и Северно-Австралийской котловин Индийского океана Амеразийской котловины Северного Ледовитого океана , тем самым начинается процесс расчленения Лавразии и Гондваны и обособления современных материков и платформ.
Конец юрского периода — время проявления позднекиммерийской фазы мезозойской складчатости в геосинклинальных поясах. В Средиземноморском поясе складкообразовательные движения проявлялись местами в начале байоса, в предкелловейское время (Крым, Кавказ), в конце юры (Альпы и др.). Но особенного размаха они достигли в Тихоокеанском поясе: в Кордильерах Северной Америки (невадийская складчатость), и Верхояно-Чукотской области (верхоянская складчатость), где сопровождались внедрением крупных гранитоидных интрузий , и завершили собой геосинклинальное развитие областей.
Органический мир Земли в юрском периоде имел типично мезозойский облик. Среди морских беспозвоночных достигают расцвета головоногие моллюски (аммониты, белемниты), широко распространены двустворчатые и брюхоногие моллюски, шестилучевые кораллы, "неправильные" морские ежи. Среди позвоночных животных в юрском периоде резко преобладают пресмыкающиеся (ящеры), которые достигают гигантских размеров (до 25-30 м) и большого разнообразия. Известны наземные травоядные и хищные ящеры (динозавры), морские плавающие (ихтиозавры, плезиозавры), летающие ящеры (птерозавры). В водных бассейнах широко распространены рыбы, в воздухе в поздней юре появляются первые (зубастые) птицы. Млекопитающие, представленные мелкими, ещё примитивными формами, мало распространены. Растительный покров суши юрского периода отличается максимумом развития голосеменных растений (цикадовые, беннетитовые, гинкговые, хвойные), а также папоротников.
От 213 до 144 млн лет назад.
К началу юрского периода гигантский сверхматерик Пангея находился в процессе активного распада. К югу от экватора все еще существовал единый обширный материк, который снова назвали Гондваной. В дальнейшем он также раскололся на части, образовавшие сегодняшние Австралию, Индию, Африку и Южную Америку. Наземные животные северного полушария уже не могли свободно перемещаться с одного материка на другой, однако они по-прежнему беспрепятственно распространялись по всему южному сверхматерику.
Вначале юрского периода климат на всей Земле был теплым и сухим. Затем, когда обильные дожди начали пропитывать влагой древние триасовые пустыни, мир вновь стал более зеленым, с более пышной растительностью. В юрском ландшафте густо росли хвощи и плауны, которые уцелели с триасового периода. Сохранились и пальмовидные беннеттиты. Кроме того, вокруг было множество гриоов. Обширные леса из семенных, обычных и древовидных папоротников, а также папо-ротникообразных саговников распространились от водоемов в глубь суши. По-прежнему были распространены хвойные леса. Кроме гинкго и араукарий, в них произрастали предки современных кипарисов, сосен и мамонтовых деревьев.
Жизнь в морях.
Когда Пангея начала раскалываться, возникли новые моря и проливы, в которых нашли прибежище новые типы животных и водорослей. Постепенно на морском дне накапливались свежие осадочные отложения. В них обосновалось множество беспозвоночных, таких, как губки и мшанки (морские циновки). В теплых и мелких морях происходили и другие важные события. Там образовались гигантские коралловые рифы, приютившие многочисленных аммонитов и новые разновидности белемнитов (давних родичей нынешних осьминогов и кальмаров).
Па суше, в озерах и реках обитало множество разных видов крокодилов, широко расселившихся по земному шару. Существовали и морские крокодилы с длинными рылами и острыми зубами для ловли рыбы. Некоторые их разновидности даже отрастили ласты вместо ног, чтобы удобнее было плавать. Хвостовые плавники позволяли им развивать в воде большую скорость, чем на суше. Появились и новые виды морских черепах. Эволюция также породила множество видов плезиозавров и ихтиозавров, соперничавших с новыми, быстроходными акулами и чрезвычайно подвижными костными рыбами.

Этот саговник - живое ископаемое. Он почти не отличается от своих родственников, произраставших на Земле в юрский период. Ныне саговники встречаются только в тропиках. Однако 200 млн лет назад они были распространены гораздо шире.
Белемниты, живые снаряды.
Белемниты были близкими родственниками современных каракатиц и кальмаров. Они имели внутренний скелет сигарообразной формы. Его основная часть, состоявшая из известкового вещества, называется ростром. В переднем конце ростра находилась полость с хрупкой многокамерной раковиной, помогавшей животному держаться на плаву. Весь этот скелет размещался внутри мягкого тела животного и служил твердым каркасом, к которому крепились его мышцы.
Твердый ростр лучше всех прочих частей тела белемнита сохраняется в ископаемом виде, и обычно именно он попадает в руки ученых. Но иногда находят и без-ростровые окаменелости. Первые подобные находки в начале XIX в. поставили в тупик многих специалистов. Они догадывались, что имеют дело с останками белемнитов, но без сопутствующего ростра эти останки выглядели довольно-таки странно. Разгадка этой тайны оказалась на редкость простой, лишь только было собрано больше данных о способе питания ихтиозавров - главных врагов белемнитов. По всей видимости, безростровые окаменелости образовывались, когда ихтиозавр, заглотав целый косяк белемнитов, изрыгал мягкие части одного из животных, в то время как его твердый внутренний скелет оставался в желудке хищника
Белемниты, подобно современным осьминогам и кальмарам, вырабатывали чернильную жидкость и использовали се для создания "дымовой завесы", когда пытались спастись от хищников. Ученые обнаружили и окаменевшие чернильные мешки белемнитов (органы, в которых хранился запас чернильной жидкости). Одному из ученых викторианской эпохи, Уильяму Бакланду, удалось даже извлечь из ископаемых чернильных мешков немного чернил, которые он использовал при иллюстрировании своей книги "Бриджуотерский трактат".

Плезиозавры, бочкообразные морские рептилии с четырьмя широкими ластами, которыми они гребли в воде, как веслами.
Склеенная подделка.
Целого ископаемого белемнита (мягкая часть плюс ростр) пока еще никому не удалось найти, хотя в 70-е гг. XX в. в Германии была предпринята довольно-таки изобретательная попытка одурачить весь научный мир с помощью искусной подделки. Цельные окаменелости, якобы добытые п одном из карьеров на юге Германии, несколько музеев приобрели но очень высокой цене, прежде чем обнаружилось, что во всех случаях известковый ростр был аккуратно приклеен к ископаемым мягким частям белемнитов!
Эта знаменитая фотография, сделанная в 1934 г. в Шотландии, была недавно объявлена подделкой. Тем не менее она пятьдесят лет подогревала энтузиазм тех, кто считал лохнесское чудовище живым плезиозавром.
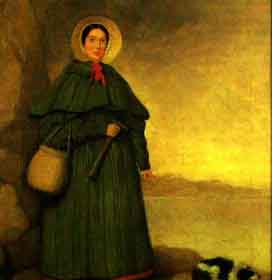
Мэри Эннинг (1799 - 1847) было всего II лет, когда она обнаружила первый ископаемый скелет ихтиозавра у Лайм-Реджис в Дороете, Англия. Впоследствии ей посчастливилось найти также первые ископаемые скелеты плезиозавра и птерозавра.
Ребенок этот мог найти
Очки, булавки, гвозди.
Но тут попались на пути
Ихтиозавра кости.
Рожденные для скорости
Первые ихтиозавры появились еще в триасе. Эти рептилии идеально приспособились к жизни в неглубоких морях юрского периода. Они обладали обтекаемым туловищем, плавниками разной величины и длинными узкими челюстями. Крупнейшие из них достигали в длину примерно 8 м, однако многие виды по размеру не превышали человека. Это были великолепные пловцы, питавшиеся главным образом рыбой, кальмарами и наутилоидеями. Хотя ихтиозавры относились к рептилиям, по их ископаемым останкам можно предположить, что они были живородящими, то есть производили на свет готовое потомство, как млекопитающие. Возможно, детеныши ихтиозавров рождались в открытом море, подобно китам.
Другая группа хищных рептилий, также широко распространенная в юрских морях, - плезиозавры. Их длинношеие разновидности обитали у поверхности моря. Здесь они охотились за косяками нсоольших рыб при помощи своих гибких шей. Короткошеие виды, так называемые плиозавры, предпочитали жизнь па больших глубинах. Они питались аммонитами и прочими моллюсками. Некоторые крупные плиозавры, по всей видимости, охотились также па менее крупных плезиозавров и ихтиозавров.

Ихтиозавры выглядели как точные копии дельфинов, за исключением формы хвоста и лишней пары плавников. Долгое время ученые считали, что у всех ископаемых ихтиозавров, попадавшихся им в руки, был поврежден хвост. В конце концов они догадались, что позвоночник этих животных имел изогнутую форму и на его конце располагался вертикальный хвостовой плавник (в отличие от горизонтальных плавников дельфинов и китов).
Жизнь в юрском воздухе.
В юрском периоде резко ускорилась эволюция насекомых, и в результате юрский ландшафт со временем наполнился нескончаемым жужжанием и потрескиванием, которые издавало множество новых видов насекомых, ползающих и летающих повсюду. Среди них были предшественники
современных муравьев, пчел, уховерток, мух и ос. Позднее, в меловом периоде, произошел новый эволюционный взрыв, когда насекомые начали "налаживать контакты" с только что появившимися цветковыми растениями.
До этого времени настоящие летающие животные встречались лишь среди насекомых, хотя попытки освоить воздушную среду наблюдались и у других существ, научившихся планировать. Теперь же в воздух поднялись целые полчища птерозавров. Это были первые и самые крупные летающие позвоночные животные. Хотя первые птерозавры появились еще в конце триаса, их подлинный "взлет" пришелся именно на юрский период. Легкие скелеты птерозавров состояли из полых костей. У первых птерозавров имелись хвосты и зубы, однако у более высокоразвитых особей эти органы исчезли, что позволило значительно уменьшить собственный вес. У некоторых ископаемых птерозавров угадывается волосяной покров. На основании этого можно предположить, что они были теплокровными.
Ученые до сих пор расходятся во мнениях относительно образа жизни птерозавров. К примеру, первоначально считалось, что птерозавры были своего рода "живыми планерами", парившими, подобно грифам, над землей в потоках поднимающегося горячего воздуха. Возможно, они даже скользили над поверхностью океана, вле-комые морскими ветрами, как современные альбатросы. Однако теперь некоторые специалисты полагают, что птерозавры могли хлопать крыльями, то есть активно летать, подобно птицам. Возможно, одни из них даже ходили по-птичьи, а другие волочили свои тела по земле или спали в местах гнездовий сородичей, повиснув вниз головой, наподобие летучих мышей.

Данные, полученные при анализе окаменевших желудков и помета (копролитов) ихтиозавров, говорят о том, что их рацион состоял в основном из рыбы и головоногих моллюсков (аммонитов, наутилоидей и кальмаров). Содержимое желудков ихтиозавров позволило сделать и еще более любопытное открытие. Маленькие твердые шипы на щупальцах кальмаров и прочих головоногих, судя по всему, доставляли ихтиозаврам немало неудобств, поскольку не переваривались и, соответственно, не могли свободно проходить через их пищеварительную систему. В результате шипы скапливались в желудке, и по ним ученым удается узнать, что данное животное съело на протяжении всей своей жизни. Так, при изучении желудка одного из ископаемых ихтиозавров выяснилось, что он проглотил по меньшей мере 1500 кальмаров!
Как птицы научились летать.
Существуют две основные теории, пытающиеся объяснить, как птицы научились летать. Одна из них утверждает, что первые полеты происходили снизу вверх. Согласно этой теории, все началось с того, что двуногие животные, предшественники птиц, разбегались и подпрыгивали высоко в воздух. Возможно, так они пытались спастись от хищников, а может, ловили насекомых. Постепенно оперенная площадь "крыльев" становилась оолыпе, прыжки, в свою очередь, удлинялись. Птица дольше не касалась земли и оставалась в воздухе. Прибавьте к этому махательные движения крыльями - и вам станет ясно, как спустя длительное время эти "пионеры воздухоплавания" научились подолгу пребывать в полете, а их крылья мало-помалу приобрели свойства, позволявшие им поддерживать тело в воздухе.
Однако существует и другая теория, противоположная, согласно которой первые полеты происходили сверху вниз, с деревьев на землю. Потенциальным "летунам" нужно было сначала забраться на значительную высоту, а уже потом броситься в воздух. В этом случае первым шагом на пути к полетам должно было стать планирование, поскольку при этом типе передвижения энергозатраты крайне незначительны - уж во всяком случае, гораздо меньшие, чем при "бегательно-пры-гательной" теории. Животному не нужно прилагать дополнительных усилий, ибо при планировании его влечет вниз сила земного тяготения.

Первого ископаемого археоптерикса обнаружили спустя два года после опубликования книги Чарлза Дарвина "Происхождение видов". Это важное открытие стало еще одним подтверждением теории Дарвина, гласившей, что эволюция происходит очень медленно и что одна группа животных порождает другую, претерпевая ряд последовательных превращений. Знаменитый ученый и близкий друг Дарвина, Томас Хаксли, предсказал существование в прошлом животного, подобного археоптериксу, еще до того, как его останки попали в руки ученых. По сути дела, Хаксли подробно описал это животное, когда оно еще не было открыто!
Ступенчатый полет.
Один ученый предложил чрезвычайно любопытную теорию. Она описывает ряд стадий, через которые должны были пройти "пионеры воздухоплавания" в ходе эволюционного процесса, превратившего их в конечном итоге в летающих животных. Согласно данной теории, когда-то одна из групп мелких рептилий, именуемых про-топтицами, перешла к древесному образу жизни. Возможно, рептилии забрались на деревья потому, что там было безопаснее, или проще добывать еду, или удобнее прятаться, спать, обустраивать гнезда. На верхушках деревьев было прохладнее, чем на земле, и у этих рептилий для лучшей теплоизоляции развились теплокровность и перьевой покров. Любые лишние длинные перья на конечностях были кстати - ведь они обеспечивали дополнительную теплоизоляцию и увеличивали площадь поверхности крылообразных "рук".
В свою очередь, мягкие, покрытые перьями передние конечности смягчали удар о землю, когда животное теряло равновесие и падало с высокого дерева. Они замедляли падение (выполняя роль парашюта), а также обеспечивали более или менее мягкую посадку, служа естественным амортизатором. Со временем эти животные стали использовать оперенные конечности в качестве протокрыльев. Дальнейший переход от пара-
тютной стадии к планирующей должен был стать вполне закономерным эволюционным шагом, после чего настала очередь последней, полетной, стадии, которой почти наверняка достиг археоптерикс.

"Ранняя" птица
Первые птицы появились на Земле ближе к концу юрского периода. Самая древняя из них, археоптерикс, больше походила на маленького пернатого динозавра, чем на птицу. У нее имелись зубы и длинный костный хвост, украшенный двумя рядами перьев. На каждом ее крыле торчало по три когтистых пальца. Одни ученые полагают, что археоптерикс пользовался своими когтистыми крыльями для лазания по деревьям, откуда он периодически слетал обратно на землю. Другие считают, что он отрывался от земли, используя порывы ветра. В процессе эволюции скелеты птиц становились все легче, а зубастые челюсти заменялись беззубым клювом. У них развилась" широкая грудина, к которой крепились мощные мышцы, необходимые для полета. Все эти изменения позволили усовершенствовать строение тела птицы, придав ему оптимальную для полета структуру.
Первой ископаемой находкой археоптерикса было единственное перо, обнаруженное в 1861 г. Вскоре в той же местности нашли цельный скелет этого животного (причем с перьями!). С тех пор обнаружено шесть окаменевших скелетов археоптерикса: одни полные, а другие лишь фрагментарные. Последняя такая находка датируется 1988 г.
Век динозавров.
Самые первые динозавры появились более 200 млн лет назад. За 140 млн лет своего существования они эволюционировали во множество самых разнообразных видов. Динозавры распространились по всем материкам и приспособились к жизни в самых различных средах обитания, хотя ни один из них не жил в норах, не лазил по деревьям, не летал и не плавал. Одни динозавры были не крупнее белки. Другие весили больше пятнадцати взрослых слонов, вместе взятых. Одни тяжело переваливались на четырех ногах. Другие бегали на двух ногах быстрее, чем олимпийские чемпионы в спринте.
65 млн лет назад псе динозавры внезапно вымерли. Однако перед тем, как исчезнуть с лица нашей планеты, они оставили нам в горных породах подробный "отчет" о своей жизни и своем времени.
Самой распространенной группой динозавров в юрском периоде были прозау-роподы. Некоторые из них развились в самых крупных наземных животных всех времен - зауроподов ("ящероногих"). Это были "жирафы" мира динозавров. Вероятно, они проводили все свое время, поедая листья с верхушек деревьев. Чтобы обеспечить жизненной энергией такое громадное тело, требовалось невероятное количество пищи. Их желудки представляли собой вместительные пищеварительные емкости, непрерывно перерабатывавшие горы растительного корма.
Позднее появилось множество разновидностей небольших быстроногих дино-
завров - так называемых гадрозавров. Это были "газели" мира динозавров. Они щипали низкорослую растительность своими роговыми клювами, а затем пережевывали ее крепкими коренными зубами.
Самым большим семейством крупных плотоядных динозавров были мегалозав-риды, или "огромные ящерицы". Мегало-заврид был чудовищем в тонну весом, с громадными острыми зубами, похожими на зубья пилы, которыми он разрывал плоть своих жертв. Судя по некоторым окаменевшим следам, пальцы его ног были обращены внутрь. Возможно, он передвигался вразвалку, подобно гигантской утке, раскачивая хвост из стороны в сторону. Мегалозавриды заселили все районы земного шара. Их ископаемые останки обнаружены в таких удаленных друг от друга местах, как Северная Америка, Испания и Мадагаскар.
Ранними видами этого семейства были, по всей видимости, относительно небольшие животные хрупкого телосложения. А более поздние мегалозавриды стали поистине двуногими чудовищами. Их задние ноги оканчивались тремя пальцами, вооруженными мощными когтями. Мускулистые передние конечности помогали при охоте на крупных растительно-ядных динозавров. Острые когти, несомненно, оставляли ужасные рваные раны в боку захваченной врасплох жертвы. Могучая мускулистая шея хищника позволяла ему со страшной силой вонзать кинжаловидные клыки глубоко в тело добычи и вырывать из нее громадные куски еще теплого мяса.

В юрском периоде на большей части земной суши разбойничали стаи аллозавров. Они, по всей видимости, представляли собой кошмарное зрелище: ведь каждый член такой стаи весил более тонны. Совместными усилиями аллозавры могли легко одолеть даже крупного зауропода.